Authors: Prof. Bhakti B. Panchal, Mrs. Geetaben Bhimani, Dr. S. K. Chawda and Dr. J.H. Rathod
Scientist, KVK, Surat,
Navsari Agriculture University, Surat, 395 007, Gujarat, India
Email: bbpanchal@nau.in
Phases of flower to fruit conversation
- Flower induction and initiation
- Flower differentiation and development
- Pollination
- Fertilization
- Fruit set and seed formation
- Growth and maturation of fruit and seed
- Fruit senescence
- An ovary wall becomes a pericarp: exocarp (fruit skin), mesocarp (fruit flesh) and endocarp (inner cell layers).
- An exocarp will develop a cuticle and may exhibit a variety of morphological features such as coarse hairs (kiwifruit) or fine hairs (peach).
- Mesocarp tissues usually represent the fleshy part of a fruit, and commonly hold chloroplasts and starch grains.
- In fleshy fruits such as berries (e.g. tomato, kiwifruit and grapes) this tissue typically comprises large parenchyma cells and contains the main vascular network.
- Endocarps are less common, but typically develop as a dense hard case around a seed, as in peach, apricot or macadamia.
- Fruits develop from organs of the flower and thus involve differentiation or redifferentiation of preexisting organs.
- Evolutionarily, floral organs represent modified leaves and so the fruit is also a modified leaf.
- Fruits serve 2 functions: to protect the seeds during development, and then to disperse the seeds following maturation.
Flower induction
- The generic term ‘fruit’ covers a wide range of structures, all supporting and protecting seeds, but where the various parts have developed from the original fertilised flower in various distinctive ways.
- Mechanistically, a fruit is a single dispersal unit which includes seeds and associated tissues, developed as a single body.
- This broad description includes structures derived from a single ovary (as in simple fruits such as apple, avocado and mango) as well as compound fruits where separate ovaries are joined (an aggregate fruit such as blackberry and cherimoya) or where separate flowers are collected into a single structure (pineapple and breadfruit).
Fertilization
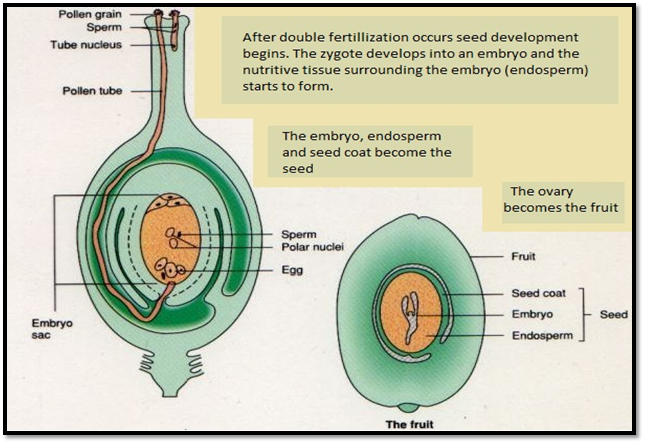
- During period of the fertilization the embryo-sac lies in a close proximity to the opening of micro pyle, into which the pollen-tube has penetrated, the separating cell-wall becomes absorbed, and the male or sperm-cells are ejected into the embryo-sac.
- Guided by the synergetic one male-cell passes into the oosphere with which it fuses, the two nuclei uniting, while the other fuses with the definitive nucleus, or, as it is also called, the endosperm nucleus.
- This is remarkable double fertilization as it has been known, although only recently discovered, has been proved to take part place in widely-separated families.
Fruit set
- Fruit set involves the decision whether to abort the ovary or proceed with fruit development. Fruit set is normally dependent on pollination.
- Pollen triggers fruit development indicating that positive signals are generated during pollination. In the absence of these signals, the flowers abscise.
- Growing pollen produces GA and application of GA can induce parthenocarpic fruit, therefore it is believed that GA is a triggering signal.
- Lagging slightly behind the growing pollen tube is a wave of increased auxin production by the style and then the ovary.
- Auxin application can also induce parthenocarpy and so it is thought that GA acts by inducing auxin production.
- An ovary must be stimulated in some way for fruit growth to occur; this is normally by pollination and fertilisation.
- This important principle was established as early that gibberellins and auxins are involved in the pollination stimulus.
- Subsequent hormone production by the fertilised ovary is critical to stimulating fruit development (Nitsch 1970).
- By implication, a suitable balance of growth regulators applied to unpollinated fruitlets can result in fruit set, and in practice gibberellins GA4 and GA7 are very effective in setting parthenocarpic (seedless) apple fruit (Dennis 1986).
- Such results confirm that growth regulators â€" alone or in combination â€" can trigger cell division in ovaries or related tissues that ultimately become fruits.
Cell division
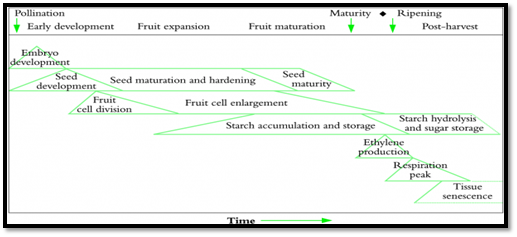
- During the first one to four weeks, flesh volume increases rapidly and embryo volume remains small.
- Growth at this time is mainly the result of cell division. In many commercial fruits (e.g. apple, kiwifruit, tomato and peach), cell division may cease a few weeks after anthesis, and fruit growth slows down, reflected as an inflection in the growth curve, and signaling an end to the first sigmoid phase.
- A second phase begins where the pericarp resumes growth and continues to enlarge until slowing for a second time as fruit mature.
- The phase of rapid cell division involves all growing parts of the fruit. This is thought to be controlled by the developing seeds.
- The number of fertilized ovules in a fruit is correlated with both the initial cell division rate and the final size of the fruit.
- Also, fruits with an uneven distribution of seeds are often lopsided. There is a correlation between cytokinin levels in developing embryos and cell division in surrounding tissues but there is no direct evidence that embryo cytokinin in fact regulates fruit cell division.
- It is difficult to reconcile the complete development of parthenocarpic fruit with the requirement of embryos for cell division except to say that parthenocarpy represents an abnormal situation.
Cell differentiation
- Patterns of cell growth and differentiation in cell layers can influence the quality of mature fruit.
- As cell size increases during development, other accompanying characteristics also change, such as cell wall thickness, differentiation of specific cell types (e.g. sclereids) and formation of cell inclusions (oil, raphides).
- Extent and distribution of airspaces are particularly important, affecting both fruit texture and physiological properties.
- This second phase in fruit growth is mainly accomplished by cell expansion in longitudinal, radial and tangential planes.
- Longitudinal growth, where cells enlarge parallel to the long axis of the fruit, will often be a big factor for development of elongate fruits such as cucumber and marrow.
- Radial growth increases diameter as in some pumpkins. Increases in cell volume during fruit growth can be considerable.
- Mature watermelons end up with some of the largest parenchyma cells in the Plant Kingdom, about 0.7 mm in diameter (Bollard 1970).
- Cell enlargement is not a uniform process. Cells in various regions of a fruit often enlarge at different rates and in different planes, so that many mature fruit show strong gradients in cell size from their surface to the centre.
- The seeds developing inside the ovary wall produce hormones.
- At first they produce cytokinins which are hormones that are exported from the seed and cause cell division in the ovary wall. This, of course, adds some thickness to the wall of the growing fruit.
- Next, the developing seeds produce gibberellic acid which is exported to the wall of the ovary and causes rapid expansion of each of the cells.
- The rate and duration of cell division varies among fruits and also among tissues within a fruit.
- Tissues made up of many small cells at maturity continue dividing while tissues composed of large cells have begun expanding.
- Cell expansion accounts for the largest increase in fruit volume, often contributing in excess of a 100 fold size increase.
- Gibberellins are also associated with fruit expansion and removal of the seeds from pea pods inhibited GA biosynthesis in the pericarp.
- Many believe that auxins from seeds regulate cell expansion of the pericarp, but auxin application does not always compensate for seed removal, and in an auxin insensitive tomato mutant, fruit growth is normal.
Seed development and Fruit growth
- Such interdependence between seed development and fruit growth shows up in final size, and apple seed numbers frequently correlate with fruit growth or with shape and size of fruit.
- As a case in point, inadequate pollination of kiwifruit results in distortion, and a curvilinear relationship emerges between seed number and fruit weight.
- A similar response is obtained when young seeds are surgically removed from immature strawberry fruits, causing a corresponding distortion in flesh development.
- Despite ample evidence that natural control of fruit shape is primarily exerted by plant hormones originating from seeds and stimulating growth to varying degrees, this is not true for all fruit.
- In banana, fertile seeds actually suppress development of the fleshy pulp. In this anomalous case, fertilisation failure allows an ovary to grow.
- In marrow, tomato and kiwifruit, ovary shape dictates spatial distribution of seeds.
- They in turn influence pericarp growth, so that fruit size and shape then become a function of initial ovary shape plus subsequent fertilisation and seed development.
Resources for fruit growth
- As fruits grow, proportions of cell wall, carbohydrate, organic acid, lipid, phospholipid and volatile (aroma) compounds change dramatically; and within each of those groups there are changes in the proportion of individual group members.
- Of these, by far the most important in practical terms is carbohydrate economy.
- Two sets of issues are at stake: (1) rate of growth, attainment of maturity and final fruit size, and (2) aroma, flavour and texture in ripe fruit. Both carry commercial implications.
- Enlarging fruit require carbohydrate to sustain cell division, enlargement and tissue specialisation.
Mineral nutrients in fruit growth
- Mineral nutrients move into the fruit most rapidly during the early stages of development at a time when xylem water flow dominates.
- As fruit approach maturity, surface to volume ratio declines, the skin becomes less permeable to water loss (sometimes through blockage of previously active stomata), and large amounts of photoassimilate are imported via phloem connections.
- As a results, a significant part of the water reaching fruit now enters through the phloem and is accompanied by photoassimilate.
- Mobile ions such as K+ and HPO42â€" are loaded into the leaf veins along with the photo-assimilate, travel in the phloem and so reach fruit over the whole growing season. In contrast, less mobile nutrients such as Ca 2+ fail to reach fruit during later stages, so that Ca 2+ concentration remains steady or even declines slightly.
- Nutrient deficiencies in fruit are relatively uncommon, except for those associated with calcium.
Maturation and ripening of fruit
- Several processes take place as fruit ripen to become edible and then senesce. They may take place while fruit are still attached or after harvest.
- Fruit are regarded as ready to harvest once they ‘mature’ because they are then capable of normal ripening off the plant.
- The combination of more cells and expanding cells leads to tremendous increase in the size of the ovary.
- As this is happening, the mother plant is producing another hormone, abscisic acid, which causes the embryo in the developing seeds to become dormant. This is adaptive because it prevents the seed from sprouting inside the warm, moist fruit.
Due to,
- Lack of pollination
- Pollination occurs but fertilization does not
- Fertilization is followed by embryo abortion
- Auxin and GA3 are well known to induce parthenocarpy. These hormonal treatment given to ovaries of unopened, unpollinated flower just before the anthesis or during the receptive period after anthesis.
- The fertilization/pollination provide other stimuli that are lacking in auxin or GA3 induce parthenocarpic fruit.
References: - Handbook of fruit production by S. Prasad & U. Kumar
- www . padlet .com/wall/ kirtivardhan
- http://plantsinaction.science.uq.edu.au/content/111-origin-fruit-tissues-and-fruit-set
- http://www.public.iastate.edu/~bot.512/lectures/seed&fruit.htm
- http://krauthammerlab.med.yale.edu Plant Physiology by S. N. Pandey and B. K. Sinha
About Author / Additional Info:
I am working as Assistant Professor(Horticulture),KVK, Surat at
Navsari Agricultural University.